Themes: Difference between revisions
mNo edit summary |
mNo edit summary |
||
| Line 1: | Line 1: | ||
===Innate immune responses at the single cell level: Experiments and modeling=== | ===Innate immune responses at the single cell level: Experiments and modeling=== | ||
[[Image:Themes-modeling_immune_responses.png|link= | [[Image:Themes-modeling_immune_responses.png|link=h|right|thumb||'''Example of pathogen recognition: LPS-induced signaling'''. LPS (lipopolysaccharide of the outer membrane of Gram-negative bacteria) is recognized by CD14 co-receptor, which transfers it to TLR4 resulting in its activation and binding of adaptor protein MyD88. Consequently, kinase TAK1 is activated and transmits signal to transcription factors p50-RelA (NF-κB) and AP-1 (early phase of ~30 min). CD14-induced endocytosis of CD14–TLR4–LPS complexes leads to TRIF-mediated activation of IRF3 and p50-RelA (late phase). Activation of transcription factors p50-RelA, IRF3 and AP1 leads to their nuclear translocation and synthesis of cytokines: TNFa, IL-1 and IFNβ that regulate via para/auto-crine signaling the second phase of innate immune responses. Transcriptional activity of p50-RelA and IRF3 is controlled by NF-κB-inducible proteins IkBα and A20 (negative feedbacks).]] | ||
Innate immunity forms the first line of defense, limiting spread of infection before the adaptive immune response is activated. In the first phase of the innate immune response, cells detect pathogens or their fragments with their membrane and cytoplasmic receptors. This leads to activation of the regulatory systems of the transcriptional factors of NF-κB, IRF3 and AP-1 families. These factors jointly regulate the activity of several hundred genes responsible for inducing inflammation, antiviral protection, proliferation, and apoptosis. In particular, they induce production and secretion of proinflammatory cytokines (among them IL-1, TNFα) as well as interferons-α and -β. These cytokines are mediators of the second phase of the cellular innate immune response in cells that did not encounter the pathogen. | Innate immunity forms the first line of defense, limiting spread of infection before the adaptive immune response is activated. In the first phase of the innate immune response, cells detect pathogens or their fragments with their membrane and cytoplasmic receptors. This leads to activation of the regulatory systems of the transcriptional factors of NF-κB, IRF3 and AP-1 families. These factors jointly regulate the activity of several hundred genes responsible for inducing inflammation, antiviral protection, proliferation, and apoptosis. In particular, they induce production and secretion of proinflammatory cytokines (among them IL-1, TNFα) as well as interferons-α and -β. These cytokines are mediators of the second phase of the cellular innate immune response in cells that did not encounter the pathogen. | ||
| Line 15: | Line 15: | ||
===Statistical methods to investigate biochemical dynamics=== | ===Statistical methods to investigate biochemical dynamics=== | ||
[[Image:Themes_experimental_design.png|link=|right| | [[Image:Themes_experimental_design.png|link=|right|thumb|'''Model-driven experimental design'''.]] | ||
Reverse-engineering of biochemical networks is largely dependent on the quality of available data. We are interested in the development of mathematical tools to help wet experimentation in obtaining more insight about the latent mechanisms that generate experimentally observed dynamics. Specifically, we incorporate available knowledge to find experimental conditions which have a potential to reveal missing information about the studied systems. We use a variety of statistical methods ranging from bayesian statistics, through model selection, to information geometry. | Reverse-engineering of biochemical networks is largely dependent on the quality of available data. We are interested in the development of mathematical tools to help wet experimentation in obtaining more insight about the latent mechanisms that generate experimentally observed dynamics. Specifically, we incorporate available knowledge to find experimental conditions which have a potential to reveal missing information about the studied systems. We use a variety of statistical methods ranging from bayesian statistics, through model selection, to information geometry. | ||
| Line 29: | Line 29: | ||
===Microfluidics for biomedical research and applications=== | ===Microfluidics for biomedical research and applications=== | ||
[[Image:Themes_microfluidics.jpg|left| | [[Image:Themes_microfluidics.jpg|left||thumb|link=http://pmbm.ippt.gov.pl/images/5/51/Themes_microfluidics.jpg|'''A microfluidic device for cell cultures'''. Inlets are connected to programmable syringe pumps which allow for precise control of the dose and duration of stimulation with, e.g., cytokines.]] | ||
Microfluidics have become an important tool in research and variety of industrial applications. Microfluidic devices enable precise operations on small volumes of liquids which can be obtain by the use of “smart” geometries or by the control of external devices. On the field of biological researches microfluidic devices can mimic physiological conditions better than other in vitro systems, and can be used for long term cell culture. They are controlled with high precision by external devices such pumps, valves and detectors, and can be easily automated. These features enable high throughput screening of the effects of different mixtures of reagents on the behaviour of cells, rendering microfluidics a powerful tool in research in cell biology and medicine. Microfluidics enables also the use of small droplets as separated chemical reactors or incubators. This approach which will be developed in the future enables to close the population of cells into small separated volume in which concentration of any reagent can be precisely adjusted. | Microfluidics have become an important tool in research and variety of industrial applications. Microfluidic devices enable precise operations on small volumes of liquids which can be obtain by the use of “smart” geometries or by the control of external devices. On the field of biological researches microfluidic devices can mimic physiological conditions better than other in vitro systems, and can be used for long term cell culture. They are controlled with high precision by external devices such pumps, valves and detectors, and can be easily automated. These features enable high throughput screening of the effects of different mixtures of reagents on the behaviour of cells, rendering microfluidics a powerful tool in research in cell biology and medicine. Microfluidics enables also the use of small droplets as separated chemical reactors or incubators. This approach which will be developed in the future enables to close the population of cells into small separated volume in which concentration of any reagent can be precisely adjusted. | ||
| Line 42: | Line 42: | ||
===Modeling of calcium signaling pathways=== | ===Modeling of calcium signaling pathways=== | ||
[[Image:Themes_equation_free_calcium_concentration.png|link=|left| | [[Image:Themes_equation_free_calcium_concentration.png|link=|left|thumb||'''A single reaction–diffusion equation for free calcium concentration'''. For a particular system of many equations, it is possible to provide a strict mathematical proof that calcium component of the traveling wave solutions tend to the traveling wave solution of the scalar equation. <!--Therefore, a system of many equations (for many kinds of buffer particles) can be replaced by a single reaction–diffusion equation, which is much easier to analyze.-->]] | ||
Calcium is one of the most important signaling messengers. Fundamental cellular processes are regulated by the influx of calcium from the extracellular matrix or its transport between intracellular compartments, e.g. cytosol ↔ mitochondria, mitochondria ↔ ER. | Calcium is one of the most important signaling messengers. Fundamental cellular processes are regulated by the influx of calcium from the extracellular matrix or its transport between intracellular compartments, e.g. cytosol ↔ mitochondria, mitochondria ↔ ER. | ||
| Line 57: | Line 57: | ||
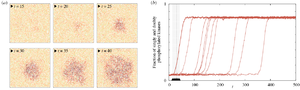
Early events in cellular communication and signaling engage proteins located in the plasma membrane. Slow diffusion therein restricts the number of interacting molecules, enhancing molecular noise, and renders the membrane a spatially organized stochastic reactor. Therefore, numerical simulation of signaling on the membrane requires methods which can account for both spatial resolution and stochastic effects, such as the kinetic Monte Carlo on the lattice. | Early events in cellular communication and signaling engage proteins located in the plasma membrane. Slow diffusion therein restricts the number of interacting molecules, enhancing molecular noise, and renders the membrane a spatially organized stochastic reactor. Therefore, numerical simulation of signaling on the membrane requires methods which can account for both spatial resolution and stochastic effects, such as the kinetic Monte Carlo on the lattice. | ||
[[Image:Themes_kmc_on_the_membrane.png|center| | [[Image:Themes_kmc_on_the_membrane.png|center||thumb|link=|'''Spontaneous initiation of activity and propagation of a traveling wave'''. ('''a''') Bistable kinase auto-phosphorylation system is activated due to a local stochastic fluctuation (orange hexagon – inactive, i.e. unphosphorylated, kinase, red – active, singly phosphorylated kinase, brown – active, doubly phosphorylated, pale green – phosphatase). Then, activity expands as a stochastic traveling wave, eventually filling whole simulated two-dimensional compartment (200×200). ('''b''') Spontaneous activation in 10 independent simulations with randomized initial conditions.]] | ||
''Selected publications'': | ''Selected publications'': | ||
Revision as of 16:15, 16 June 2017
Innate immune responses at the single cell level: Experiments and modeling
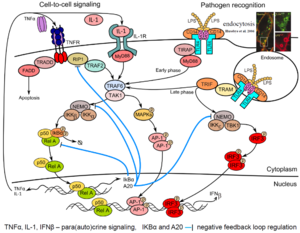
Innate immunity forms the first line of defense, limiting spread of infection before the adaptive immune response is activated. In the first phase of the innate immune response, cells detect pathogens or their fragments with their membrane and cytoplasmic receptors. This leads to activation of the regulatory systems of the transcriptional factors of NF-κB, IRF3 and AP-1 families. These factors jointly regulate the activity of several hundred genes responsible for inducing inflammation, antiviral protection, proliferation, and apoptosis. In particular, they induce production and secretion of proinflammatory cytokines (among them IL-1, TNFα) as well as interferons-α and -β. These cytokines are mediators of the second phase of the cellular innate immune response in cells that did not encounter the pathogen.
Selected publications:
- Barua D, Hlavacek WS, Lipniacki T. A computational model for early events in B cell antigen receptor signaling: analysis of the roles of Lyn and Fyn, J Immunol 189:646-658 (2012) PubMed CrossRef | PDF-ms
- Hat B, Kaźmierczak B, Lipniacki T. B cell activation triggered by the formation of the small receptor cluster: a computational study, PLoS Comput Biol 7(10):e1000448 (2011) PubMed CrossRef | PDF
- Tay S, Hughey J, Lee T, Lipniacki T, Covert M, Quake S. Single-cell NF-κB dynamics reveal digital activation and analogue information processing, Nature 466:267-271 (2010) PubMed CrossRef | PDF Code
Statistical methods to investigate biochemical dynamics

Reverse-engineering of biochemical networks is largely dependent on the quality of available data. We are interested in the development of mathematical tools to help wet experimentation in obtaining more insight about the latent mechanisms that generate experimentally observed dynamics. Specifically, we incorporate available knowledge to find experimental conditions which have a potential to reveal missing information about the studied systems. We use a variety of statistical methods ranging from bayesian statistics, through model selection, to information geometry.
Selected publications:
- Liepe J, Filippi S, Komorowski M, Stumpf MPH. Maximising the information content of experiments in systems biology, PLOS Comput Biol 9(1):e1002888 (2013) PubMed CrossRef
- Komorowski M, Costa MJ, Rand DA, Stumpf MPH. Sensitivity, robustness, and identifiability in stochastic chemical kinetics models, Proc Natl Acad Sci USA 108(21):8645-50 (2011) PubMed CrossRef
- Komorowski M, Finkenstädt B, Harper CV, Rand DA. Bayesian inference of biochemical kinetic parameters using the linear noise approximation, BMC Bioinformatics 10:343 (2009) PubMed CrossRef
Microfluidics for biomedical research and applications

Microfluidics have become an important tool in research and variety of industrial applications. Microfluidic devices enable precise operations on small volumes of liquids which can be obtain by the use of “smart” geometries or by the control of external devices. On the field of biological researches microfluidic devices can mimic physiological conditions better than other in vitro systems, and can be used for long term cell culture. They are controlled with high precision by external devices such pumps, valves and detectors, and can be easily automated. These features enable high throughput screening of the effects of different mixtures of reagents on the behaviour of cells, rendering microfluidics a powerful tool in research in cell biology and medicine. Microfluidics enables also the use of small droplets as separated chemical reactors or incubators. This approach which will be developed in the future enables to close the population of cells into small separated volume in which concentration of any reagent can be precisely adjusted.
Selected publications:
- Węgrzyn J, Samborski A, Reissig L, Korczyk PM, Błoński S, Garstecki P. Microfluidic architectures for efficient generation of chemistry gradations in droplets, Microfluid Nanofluid 14(1-2):235–245 (2013, e-pub 2012) CrossRef | PDF
- Jakiela S, Korczyk PM, Makulska S, Cybulski O, Garstecki P. Discontinuous transition in a laminar fluid flow: A change of flow topolgy inside a droplet moving in a micron-size channel, Phys Rev Lett 108(13):134501 (2012) PubMed CrossRef
- Korczyk PM, Cybulski O, Makulska S, Garstecki P. Effects of unsteadiness of the rates of flow on the dynamics of formation of droplets in microfluidic systems, Lab Chip 11:173-175 (2011) PubMed CrossRef
Modeling of calcium signaling pathways
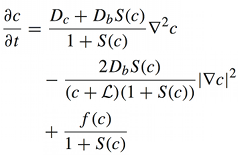
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Calcium is one of the most important signaling messengers. Fundamental cellular processes are regulated by the influx of calcium from the extracellular matrix or its transport between intracellular compartments, e.g. cytosol ↔ mitochondria, mitochondria ↔ ER.
Selected publications:
- Kaźmierczak B, Peradzyński Z. Calcium waves with fast buffers and mechanical effects, J Math Biol 62:1-38 (2011, online 2010) PubMed CrossRef
- Kaźmierczak B, Volpert V. Calcium waves in systems with immobile buffers as a limit of waves for systems with non zero diffusion, Nonlinearity 21:71 (2008) CrossRef
- Kaźmierczak B, Volpert V. Travelling calcium waves in systems with non diffusing buffers, Math Mod Meth Appl Sci 18(6):883-912 (2008) CrossRef
Kinetic Monte Carlo simulations of biochemical reactions on the plasma membrane
Early events in cellular communication and signaling engage proteins located in the plasma membrane. Slow diffusion therein restricts the number of interacting molecules, enhancing molecular noise, and renders the membrane a spatially organized stochastic reactor. Therefore, numerical simulation of signaling on the membrane requires methods which can account for both spatial resolution and stochastic effects, such as the kinetic Monte Carlo on the lattice.
{kind=link}
Selected publications:
- Kochańczyk M, Jaruszewicz J, Lipniacki T. Stochastic transitions in a bistable reaction system on the membrane, J R Soc Interface 10(84):20130151 (2013) PubMed CrossRef | PDF Supp-PDF Supp-movies
- Żuk PJ, Kochańczyk M, Jaruszewicz J, Bednorz W, Lipniacki T. Dynamics of a stochastic spatially extended system predicted by comparing deterministic and stochastic attractors of the corresponding birth-death process, Phys Biol 5(9):055002 (2012) PubMed CrossRef | PDF-ms
- Szymańska P, Kochańczyk M, Miększ J, Lipniacki T. Effective reaction rates in diffusion-limited phosphorylation–dephosphorylation cycles, Phys Rev E 91:022702 (2015) CrossRef | PDF-ms